What components of sound are used for hearing?
There are a wide variety of ear structures in terrestrial and aquatic vertebrates and invertebrates. Ears in marine invertebrates and fishes evolved prior to ears in land animals whereas ears of marine mammals evolved from those of land mammals that returned to water and therefore readapted to hearing in water. Scientists investigate how ears of each aquatic group work and how they may differ from land based counterparts.
Sound waves compress and expand the medium as they propagate through it. This represents the pressure component of sound. The particles of the medium vibrate back and forth along the direction of propagation (particle motion).
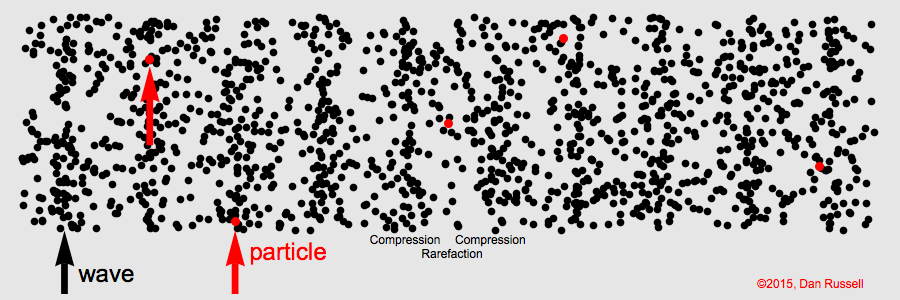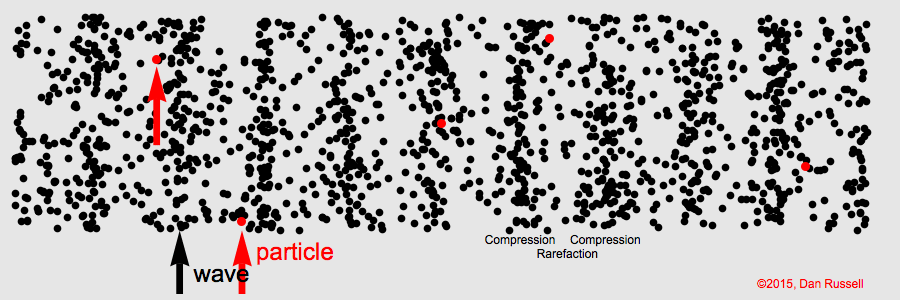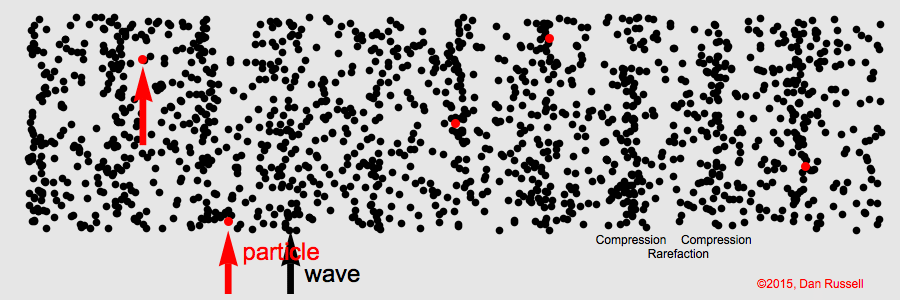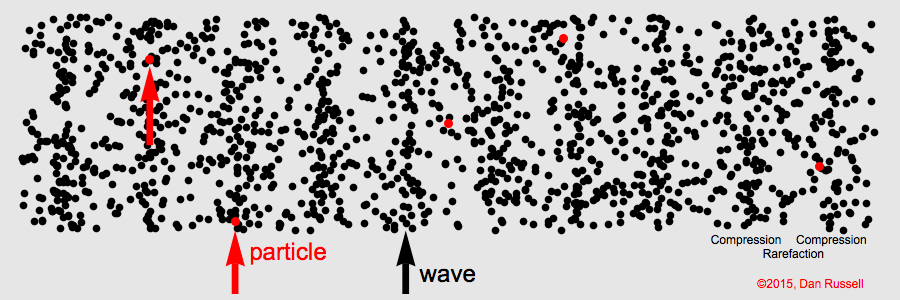
Sound is a longitudinal wave. Animation credit: Dan Russell, Grad. Prog. Acoustics, Penn State, https://www.acs.psu.edu/drussell/Demos/waves/wavemotion.html
The energy transmitted by a sound wave is comprised of both pressure (potential energy due to the acoustic impedance) and particle velocity (kinetic energy due to the movement of the particles) components. Acoustic intensity is defined as the amount of energy transmitted through a unit area per unit of time in the direction in which the sound wave is traveling; it is a measure of acoustic power. Acoustic intensity is the product of the pressure and particle velocity of the sound wave:
$latex \overrightarrow{I} = p \overrightarrow{u} &s=2$
intensity ( $latex \overrightarrow{I} &s=1$ ), pressure (p), particle velocity ( $latex \overrightarrow{u} &s=1$ )
The sound is influenced by the medium through which it moves. The ratio of acoustic pressure (p) to the associated particle speed in a medium (u) is called the specific acoustic impedance (z). This quantity is similar in concept to the idea of electrical resistance.
z = p/u
A similar concept, acoustic impedance (Z), depends on the medium and the dimensions of the enclosed volume in which the medium is contained. The acoustic impedance is defined as the ratio of the average sound pressure across a specific surface to the volume velocity (U):
Z = p/U
Animals can detect these different aspects of sound. Some species, such as marine fishes and some invertebrates are thought to use particle motion; others, such as marine mammals, are thought to detect intensity and therefore use both pressure and velocity components.
Particle motion is described by displacement, velocity, and acceleration. All fishes detect particle motion, while some also detect pressure and possibly intensity. (For more on fish hearing see: How do fish hear sounds?) While there are few studies of hearing by marine invertebrates, available data suggest that some invertebrates detect particle motion. (For more on invertebrate hearing see: How do marine invertebrates detect sounds?)
Most fish and invertebrate tissues have a density that is approximately the same as water and move with the impinging sound wave. Thus, they do not detect sound pressure. Most bony fishes have solid calcified masses called otoliths (literally, “ear stones”). More primitive fish species (such as sturgeon and paddlefish) and elasmobranchs (sharks and rays) have multiple, separate crystals, called otoconia, held together by gelatinous material. The otoliths or otoconia contact the stereociliary bundles of the hair cells. Because the density of otoliths and otoconia is higher than water, they move with a different amplitude and phase than the rest of the fish’s tissues, causing the stereociliary bundles of the hair cells to bend. This results in a neural signal being sent to the brain to convey the detected particle motion component of sound.
Fish hearing short: Otoliths. Video describing fish otoliths and how they work. Video by Discovery of Sound in the Sea.
Some cephalopods (octopus and squid) and decapod crustaceans (lobster, shrimp, and crab) sense the particle motion component of sound[1]Packard, A., Karlsen, H. E., & Sand, O. (1990). Low frequency hearing in cephalopods. Journal of Comparative Physiology A, 166(4). https://doi.org/10.1007/BF00192020[2]Budelmann, B. U. (1992). Hearing in Nonarthropod Invertebrates. In D. B. Webster, A. N. Popper, & R. R. Fay (Eds.), The Evolutionary Biology of Hearing (pp. 141–155). New York, NY: Springer New York. https://doi.org/10.1007/978-1-4612-2784-7_10[3]Budelmann, B. U., & Williamson, R. (1994). Directional sensitivity of hair cell afferents in the octopus statocyst. The Journal of Experimental Biology, 187, 245–259.. It has been suggested that this is done in the statocyst, which also detects gravity and acceleration of the animal. Similar to otoliths, statocysts contain a dense material, that may be sand grains or other materials, that moves differently than the rest of the body.
Most marine crustaceans (e.g., lobster, crabs, and shrimp) also have an extensive array of sensory hairs covering their bodies. Water- or substrate-borne vibrations associated with changes in acceleration, hydrodynamic flow, and/or sound, may stimulate these sensory hairs.
Some bony fishes are also able to detect the pressure component of sound using a gas-filled structure, the swim bladder, located in the abdominal cavity. Because gas is much more compressible than water, the swim bladder will expand and contract with the pressure changes that occur within a sound field. This pulsation reradiates the received sound pressure. This reradiated signal travels through fish tissues to the ear and may stimulate the ear. How well the signal from the swim bladder is detected at the ear depends on how much the signal is attenuated between the swim bladder and the ear. If the fish has adaptions to the swim bladder that bring it closer to the ear, then the signal is not attenuated very much. In these cases, the fish is able to detect sound pressure as well as particle motion [4]Schulz-Mirbach, T., Ladich, F., Mittone, A., Olbinado, M., Bravin, A., Maiditsch, I. P., Melzer, R. R., Krysl, P., & Heß, M. (2020). Auditory chain reaction: Effects of sound pressure and particle motion on auditory structures in fishes. PLOS ONE, 15(3), e0230578. https://doi.org/10.1371/journal.pone.0230578.[5]Popper, A. N., & Hawkins, A. D. (2018). The importance of particle motion to fishes and invertebrates. The Journal of the Acoustical Society of America, 143(1), 470–488. https://doi.org/10.1121/1.5021594..

Microtomographic (microCT) imaging showing the complex swim bladder-inner ear connection in the Asian cichlid (Etroplus maculatus) in which a bipartite swim bladder extension contacts the upper as well as the lower parts of each inner ear. Images adapted from Schulz-Mirbach, T., Heß, M., Metscher, B.D. et al. A unique swim bladder-inner ear connection in a teleost fish revealed by a combined high-resolution microtomographic and three-dimensional histological study. BMC Biol 11, 75 (2013). https://doi.org/10.1186/1741-7007-11-75.
One group of species, the otophysans (which include carps, minnows, catfishes, and characins) have a unique mechanical coupling, the Weberian ossicles, connecting the swim bladder and inner ear. The Weberian ossicles are a series of modified bones of the vertebral column (the first few vertebrae of the backbone). They are thought to move in response to sound stimuli that cause movements of the wall of the swim bladder and carry the signal to the inner ear, thereby directly causing movement of the otoliths, and stimulating hearing [6]Schulz-Mirbach, T., Ladich, F., Mittone, A., Olbinado, M., Bravin, A., Maiditsch, I. P., Melzer, R. R., Krysl, P., & Heß, M. (2020). Auditory chain reaction: Effects of sound pressure and particle motion on auditory structures in fishes. PLOS ONE, 15(3), e0230578. https://doi.org/10.1371/journal.pone.0230578.. As a result of having the Weberian ossicles, most otophysans hear less intense sounds and can hear to higher frequencies than most other fish species[7] Popper, A. N., and A.D. Hawkins. (2021). “Hearing,” in The Physiology of Fishes, edited by S. Currie, and D. H. Evans (CRC Press, Boca Raton, FL). Indeed, goldfish hear up to 3 kHz with best hearing from 500-800Hz.
It has been suggested that marine mammals may detect sound pressure only but the majority of research indicates that they, like land mammals, operate primarily as intensity detectors[8]Finneran, J. J., Carder, D. A., & Ridgway, S. H. (2002). Low-frequency acoustic pressure, velocity, and intensity thresholds in a bottlenose dolphin ( Tursiops truncatus ) and white whale ( Delphinapterus leucas ). The Journal of the Acoustical Society of America, 111(1), 447–456. https://doi.org/10.1121/1.1423925. It is difficult both in air and in water to measure intensity per se, therefore differentially testing sound pressure reception has not yet been done for aquatic mammals[9]Lynch, T. J., Nedzelnitsky, V., & Peake, W. T. (1982). Input impedance of the cochlea in cat. The Journal of the Acoustical Society of America, 72(1), 108–130. https://doi.org/10.1121/1.387995[10]Peake, W. T., Rosowski, J. J., & Lynch, T. J. (1992). Middle-ear transmission: Acoustic versus ossicular coupling in cat and human. Hearing Research, 57(2), 245–268. https://doi.org/10.1016/0378-5955(92)90155-G[11]Brundin, L., & Russell, I. (1994). Tuned phasic and tonic motile responses of isolated outer hair cells to direct mechanical stimulation of the cell body. Hearing Research, 73(1), 35–45. https://doi.org/10.1016/0378-5955(94)90280-1. (See How do marine mammals hear?)
Measuring Sound Pressure and Particle Velocity
The most common device for measuring underwater sound is a hydrophone. Hydrophones measure the pressure (amplitude) of a sound wave, rather than its intensity. Hydrophones are not sensitive to particle velocity. They estimate sound intensity level from the measured sound pressure by assuming that the acoustic field can be modeled as a propagating plane wave. In this case, particle velocity is assumed to be proportional to and in phase with acoustic pressure. (For more detail, see: Introduction to Decibels)
Geophones measure particle velocity, but are more commonly used to measure seismic waves than underwater sound waves. Vector sensors measure particle velocity or acceleration together with pressure to provide direct measurements of intensity, including the direction in which the sound wave is traveling. Vector sensors are used in DIFAR (Directional Frequency Analysis and Recording) sonobuoys, for example.
Additional Links on DOSITS
- Animals and Sound > How do fish hear?
- Animals and Sound > How do marine mammals hear?
- Science of Sound > What is intensity? (Advanced Topic)
- Science of Sound > What is sound?
- Science of Sound > How does sound in air differ from sound in water?
- Science of Sound > Introduction to Decibels (Advanced Topic)
- Technology Gallery > Geophone
- Technology Gallery > Vector Sensors
- Technology Gallery > Hydrophone/Receiver
- Technology Gallery > Sonobuoys
Additional Resources
- Acoustics and Vibration Animations by Dr. Daniel A. Russell: Longitudinal and Transverse Wave Motion
- Yost, W. A. (2020). Dick Fay and Goldfish. Acoustics Today, 16(3), 53. https://doi.org/10.1121/AT.2020.16.3.53.
References
- Fay, R. (1984). The goldfish ear codes the axis of acoustic particle motion in three dimensions. Science, 225(4665), 951–954. https://doi.org/10.1126/science.6474161.
- Hawkins, A. D., Hazelwood, R. A., Popper, A. N., & Macey, P. C. (2021). Substrate vibrations and their potential effects upon fishes and invertebrates. The Journal of the Acoustical Society of America, 149(4), 2782–2790. https://doi.org/10.1121/10.0004773.
-
Hawkins, A. D., & Popper, A. N. (2020). Sound detection by Atlantic cod: An overview. The Journal of the Acoustical Society of America, 148(5), 3027–3041. https://doi.org/10.1121/10.0002363.Nedelec, S. L., Campbell, J., Radford, A. N., Simpson, S. D., & Merchant, N. D. (2016). Particle motion: The missing link in underwater acoustic ecology. Methods in Ecology and Evolution, 7(7), 836–842. https://doi.org/10.1111/2041-210X.12544.
-
Popper, A. N., & Hawkins, A. D. (2018). The importance of particle motion to fishes and invertebrates. The Journal of the Acoustical Society of America, 143(1), 470–488. https://doi.org/10.1121/1.5021594.
-
Schulz-Mirbach, T., & Ladich, F. (2016). Diversity of Inner Ears in Fishes: Possible Contribution Towards Hearing Improvements and Evolutionary Considerations. In J. A. Sisneros (Ed.), Fish Hearing and Bioacoustics (Vol. 877, pp. 341–391). Springer International Publishing. https://doi.org/10.1007/978-3-319-21059-9_16.
- Wartzok, D., & Ketten, D. R. (1999). Marine Mammal Sensory Systems. In J. E. I. Reynolds & S. E. Rommel (Eds.), Biology of Marine Mammals (pp. 117–175). Washington D.C.: Smithsonian Institution Press.
- Webb, J. F., Fay, R. R., & Popper, A. N. (Eds.). (2008). Fish bioacoustics. New York: Springer.
- Webb, J. F., Herman, J. L., Woods, C. F., & Ketten, D. R. (2010). The ears of butterflyfishes (Chaetodontidae): ‘Hearing generalists’’ on noisy coral reefs?’ Journal of Fish Biology, 77(6), 1406–1423. https://doi.org/10.1111/j.1095-8649.2010.02765.x.
Cited References
| ⇡1 | Packard, A., Karlsen, H. E., & Sand, O. (1990). Low frequency hearing in cephalopods. Journal of Comparative Physiology A, 166(4). https://doi.org/10.1007/BF00192020 |
|---|---|
| ⇡2 | Budelmann, B. U. (1992). Hearing in Nonarthropod Invertebrates. In D. B. Webster, A. N. Popper, & R. R. Fay (Eds.), The Evolutionary Biology of Hearing (pp. 141–155). New York, NY: Springer New York. https://doi.org/10.1007/978-1-4612-2784-7_10 |
| ⇡3 | Budelmann, B. U., & Williamson, R. (1994). Directional sensitivity of hair cell afferents in the octopus statocyst. The Journal of Experimental Biology, 187, 245–259. |
| ⇡4, ⇡6 | Schulz-Mirbach, T., Ladich, F., Mittone, A., Olbinado, M., Bravin, A., Maiditsch, I. P., Melzer, R. R., Krysl, P., & Heß, M. (2020). Auditory chain reaction: Effects of sound pressure and particle motion on auditory structures in fishes. PLOS ONE, 15(3), e0230578. https://doi.org/10.1371/journal.pone.0230578. |
| ⇡5 | Popper, A. N., & Hawkins, A. D. (2018). The importance of particle motion to fishes and invertebrates. The Journal of the Acoustical Society of America, 143(1), 470–488. https://doi.org/10.1121/1.5021594. |
| ⇡7 | Popper, A. N., and A.D. Hawkins. (2021). “Hearing,” in The Physiology of Fishes, edited by S. Currie, and D. H. Evans (CRC Press, Boca Raton, FL) |
| ⇡8 | Finneran, J. J., Carder, D. A., & Ridgway, S. H. (2002). Low-frequency acoustic pressure, velocity, and intensity thresholds in a bottlenose dolphin ( Tursiops truncatus ) and white whale ( Delphinapterus leucas ). The Journal of the Acoustical Society of America, 111(1), 447–456. https://doi.org/10.1121/1.1423925 |
| ⇡9 | Lynch, T. J., Nedzelnitsky, V., & Peake, W. T. (1982). Input impedance of the cochlea in cat. The Journal of the Acoustical Society of America, 72(1), 108–130. https://doi.org/10.1121/1.387995 |
| ⇡10 | Peake, W. T., Rosowski, J. J., & Lynch, T. J. (1992). Middle-ear transmission: Acoustic versus ossicular coupling in cat and human. Hearing Research, 57(2), 245–268. https://doi.org/10.1016/0378-5955(92)90155-G |
| ⇡11 | Brundin, L., & Russell, I. (1994). Tuned phasic and tonic motile responses of isolated outer hair cells to direct mechanical stimulation of the cell body. Hearing Research, 73(1), 35–45. https://doi.org/10.1016/0378-5955(94)90280-1 |